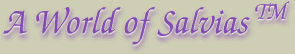
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
|
A typical Salvia greggii was chosen as the model for these illustrations. All the features shown are present in all Salvia flowers, except for the invagination found in S. microphylla and its hybrids. Other possible features for other species are not covered. Flower
Stem Structures
|
Flower Stem Structures
|
This is a mature spike of a Salvia greggii. It shows the most common features found in Salvia flowers. The floral leaves (a) are only slightly reduced in this form from typical leaves. In many broadleaf sages, they are often reduced in size. Further, they can develop progressively into bracts by taking on some of their features like vein patterns and shape. For an example of this, see the image of S. wagneriana. The petiole (leaf stem) is not shown on this image. The calyx (c) is the cup-like structure that holds the flower (d, corolla) and ovary, connected to the floral stem by a stalk (b, the pedicel). In some sages, the calyx becomes swollen (accrescent) after the flower drops, most frequently if the ovary has been successfully fertilized. Abscission is the process of the deliberate separation of a part of a plant like a flower, fruit, or old leaf from the plant. On most sages, non-fertile calyxes fall off in a few days after the flowers. When calyxes persist and are brightly colored, they add significantly to the floral display. Examples are S. splendens, S. mexicana, and S. regla. There is often a ring of hairs (annulus) towards the mouth that holds in the ripening seed. Immature calyxes often have the mouth flattened shut, and calyxes with ripening seed (e) often open up as they dry and brown up. Actually, seeds in the mint family are really achenes, or one-seeded fruit. They are also referred to as nutlets. |
![]()
Flower Tissue Parts
|
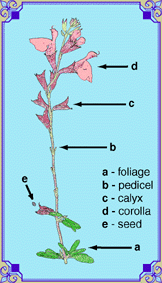
This illustration shows the most visible parts of the sage flower. All sages have these parts, in different ratios. This particular form, typical of S. greggii, S. coccinea, S. urica, and S. farinacea, is one of the most common, with no one part dominant over the others. In the mint family (Labiatae or Lamiaceae), all flowers are assymetric. The greater group of plants to which the mints belong are bell-shaped (campanulate), and they usually have five-fold symmetry. On some sports of sages, the stem come to an abrupt halt with a terminal flower (a determinate stem). This peloric flower has the five-fold symmetry of its distant cousins. For an example, see the Huntington Botanical Garden selection of S. iodantha. Another example of a peloric flower is the solitary deep purple flower in the center of the wild carrot umbel. In mints, the bell remains as the corolla tube (a). It is often swollen in the middle to facilitate the lever process of the stamen. One of the petals has become the upper lip, or hood (b). This structure has developed to protect the style and anther until they have matured. The longest hairs on sage flowers are usually found here The remaining four petals show up as the rounded prominance made from the four lobes on the lower lip (c) or palate. The lower two lobes (d) are the most obvious. The hairs on these parts are much finer. Most of the color on sage flowers are in these hairs. The geometry of these hairs may have a profound effect on the perceived color of the flower due to reflective angles, refraction of light, and other optical effects. The main differences between different sage flowers are the variations in the geometries of the different parts. Some have very small lower lips, and others have elongated corolla tubes. |
![]()
Sexual Organs - Pistil and Stamen
![]()
Secondary Floral Features
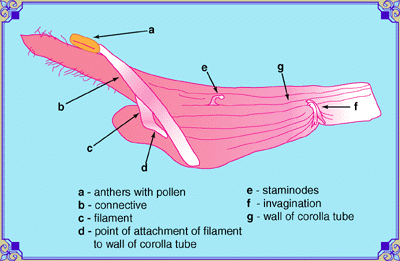
|
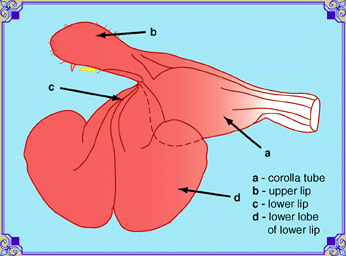
There are other features not normally noticed on Salvia flowers that can be useful for the description of the species. These include the staminodes (e), the vestiges of the second pair of stamens. There are two stamens in the genus Salvia, not four, as there are in most other members of the mint family. Many people think Stachys coccinea is a Salvia, but a quick inspection shows four anthers. In S. microphylla, there are a pair of invaginations or papillae that protrude at the constriction near the base of the corolla. Imagine that the tissue of the flower has the consistancy of silly putty, and you have poked your finger in it, then withdrawn to leave a horn-like, curved, sealed tube or a pimple. In hybrids with S. greggii, the structure is still there, but reduced. Their function is supposed to guide the tongue of the pollinator to the nectary. In this image, the lever made by the filament (c) to raise the connective (b) from the corolla tube wall (g) can be seen more clearly. |
![]()
Seed Development in the Ovary
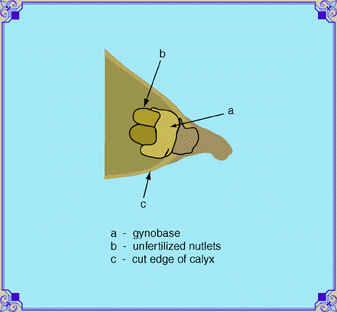
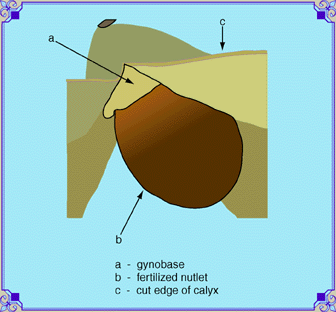
| The gynobase (seed base, a) support the ovules that
form the future nutlets (achenes, b).
The base is attached to the end of the floral stem (pedicel).
These parts and the base of the calyx constitute the
ovary.
There are four ovules attached to the base in a two by two array. The pistil is attached to the gynobase in the midst of the four seeds. The sperm tube continues until it reaches the unfertilized seed through the point of attachment. It passes through to the ovule (unfertilized seed), leaving a microscopic hole (micropyle),. through which the germinating seed sends its emerging root. The scar where the seed was attached is called the hilum. Seeds are dark brown or black when they are ripe, and will break off easily at this stage. If you want to collect seed on sages, pinch the mouth of the calyx shut before pulling it off. |
![]()
![]()
Links to Other World of Salvia Pages:
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()